Co-transport
This is also called
secondary active transport or
indirect active transport.
It does not directly use ATP as an energy source, but rather it causes the
movement of one substance together with another which has already established an electrochemical potential gradient
after being transported using ATP as above. It also requires a specific
carrier protein (cotransporter) which can accommodate
both substances, and this movement is facilitated diffusion.
Thus, by one substance passing back across the membrane down its concentration gradient, it allows the passage of another substance across the membrane. In some cases, both substances pass across the membrane in the same direction; in others they move in opposite directions.
Co-transport of sodium ions and glucose
- both going in the same direction

In the absorption of sodium ions and glucose by epithelial cells lining the villi within the mammalian ileum (
section of inner membrane shown above), these cells have sodium ions actively pumped out of them on the outer surface membrane, so that the concentration of Na
+ ions inside the cell is lower than in the lumen of the gut (the space containing food digestion products).
In other words, it maintains a diffusion gradient for Na
+ ions from the lumen into the gut lining cells, and this provides a driving force for the entry of both sodium ions and glucose from the digested food via the symporter protein.
(
More details opposite)
Co-transport of sodium ions and calcium ions
- each going in opposite directions

This co-transport relies on the higher concentration of sodium ions outside the cell (above the membrane), thus an influx of sodium ions powers the exit of the calcium ions.
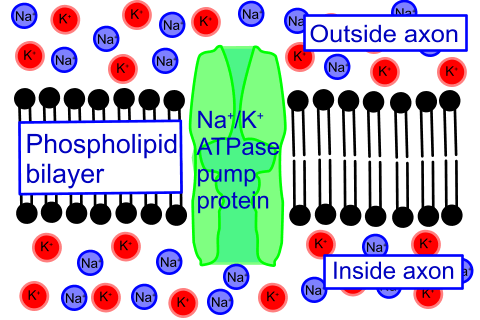
Secondary active transport may also be referred to as
ion-coupled transport, and it often relies on sodium ions Na
+ or protons H
+ which are accumulated by ATP-driven active transport pumps..
There are two classes of secondary transfer, named according to the carrier protein and the direction of transport of the two substances involved.
In each case, one substance powers the exchange by moving down its potential energy gradient, previously established by an ATP-powered pump.
Symporters
Both substances are transported across the membrane in the same direction.
Example:
Co-transport mechanism for the absorption of glucose into the blood by a cell lining the ileum
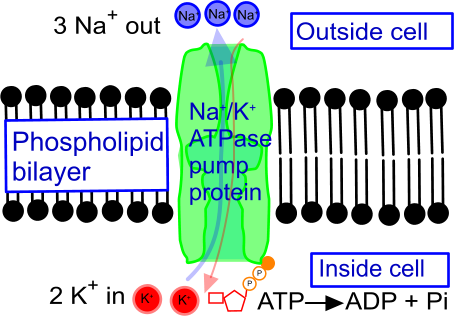
 Suggest a name for the structures labelled with letters:
Suggest a name for the structures labelled with letters:
A:
co-transport protein/glucose symporter/SGLT1
B:
Na+/K+ ATPase pump
C:
carrier protein/glucose permease/GLUT2
In the absorption of glucose into cells lining the villi within the ileum (small intestine),
the
glucose symporter (SGLT1) co-transports one glucose molecule into the cell for every two sodium ions it imports into the cell. The inward movement of sodium ions is assured by a sodium-potassium pump (powered by ATP) at the
other end of the cell which reduces the internal concentration of sodium ions. Glucose (together with sodium ions) then diffuses out of the cell via a channel protein (GLUT2) - this is
facilitated diffusion - and into a blood capillary on the outside of the cell.
What causes glucose to diffuse in this direction?
>
Movement down a concentration gradient - glucose builds up in the cell as it is brought in by the symporter, and the flow in the capillary ensures a constant lower concentration at this end of the cell.
There are also a number of membrane-bound symporters that co-transport amino acids together with sodium ions into the epithelium lining the ileum.
Which of the structures (A, B and C) in the diagram opposite would also participate in the uptake of an amino acid?
>
Only B - A and C would be specific for glucose
See the photomicrograph below showing brush border.
A different symporter (SGLT2) is also located in the proximal tubule in the kidney nephrons where it is responsible for the selective reabsorption of glucose into the blood.
Antiporters
Here the two substances move across the membrane in opposite directions.
Example:
The
sodium-calcium ion exchanger or antiporter, which allows three sodium ions into the cell to transport one calcium out.
This is important in neurones and cardiac muscle cells as it acts quickly to reduce the cytoplasmic calcium concentration after activity.
[This is in fact additional to Plasma membrane Ca
2+ ATPase (PMCA), which is a transport protein in the plasma membrane of all eukaryotic cells that also serves to maintain low concentrations of calcium (Ca
2+) within the cells.]
Nitrate ion uptake from soil water and passage across the root
There are said to be 2 pathways though which water and solutes can pass between cells: the symplast - the cytoplasm within cells, where the enzyme nitrate reductase can convert nitrate ions to nitrite - and the apoplast - basically the inactive cell walls which provide a less selective route.
Nitrate ions (NO
3-) enter the symplast (cytoplasm) of a root hair cell by means of an H
+/NO
3- symporter and remain in the symplast as they travel inwards from cell to cell toward the root xylem, moving by means of plasmodesmata - cytoplasmic strands linking cells directly. Once in the xylem, the NO
3- ion travels upwards towards the leaf in the transpiration stream.
Uptake of nitrate is probably best seen as secondary active transport, and the activity is not confined to root hair cells, as the nitrate reductase activity creates a concentration gradient across the root.

![An electron micrograph of the plasma membrane (of Chara corallina (cropped from [1]). To fix the membrane for electronmicroscopy the material was crosslinked using OsO 4](https://www.researchgate.net/profile/Hans_Coster/publication/235368403/figure/fig1/AS:299822390235137@1448494636506/An-electron-micrograph-of-the-plasma-membrane-of-Chara-corallina-cropped-from-1-To.png)

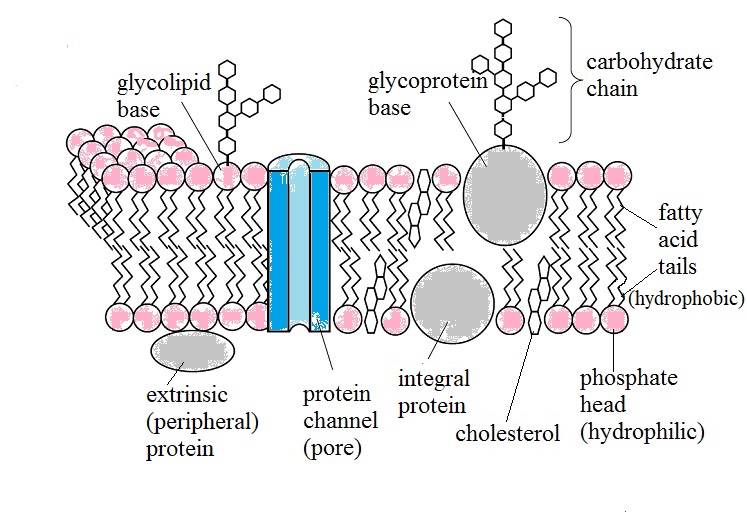
 A nice mosaic with a Biological feel, but it is rather static
A nice mosaic with a Biological feel, but it is rather static
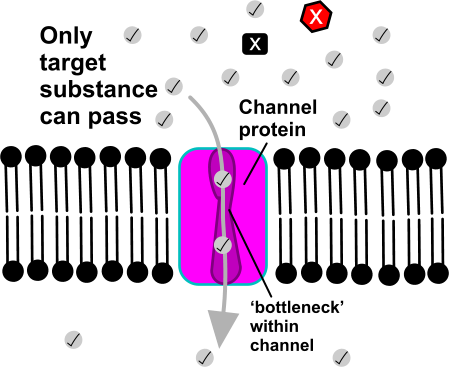

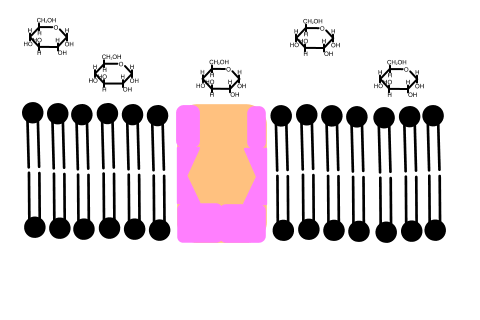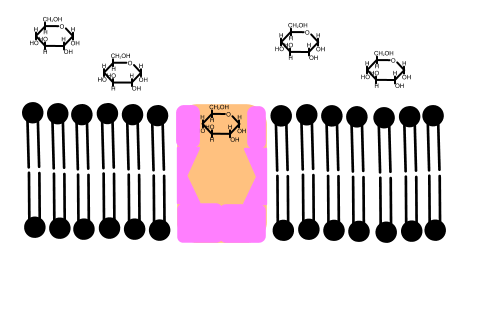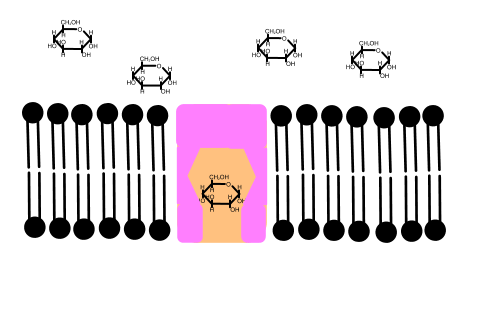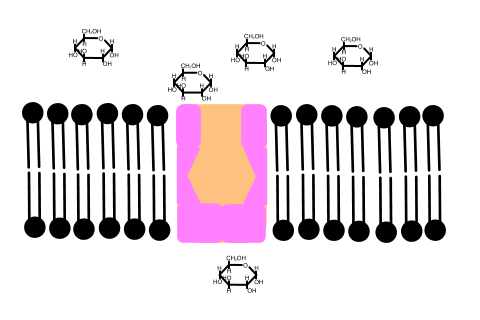
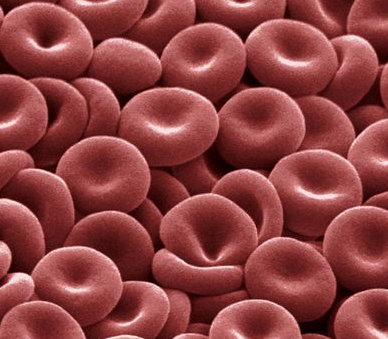
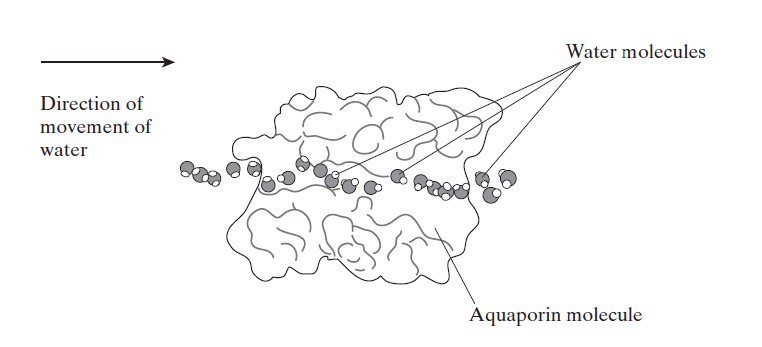 In 1992 Peter Agre showed the molecular structure of a water channel called aquaporin in red blood cells and kidney tubules and he received the Nobel prize for this in 2003 . Other versions have been discovered in other cells, including plants.
In 1992 Peter Agre showed the molecular structure of a water channel called aquaporin in red blood cells and kidney tubules and he received the Nobel prize for this in 2003 . Other versions have been discovered in other cells, including plants.
 Cells lining the small intestine, showing a brush border
Cells lining the small intestine, showing a brush border
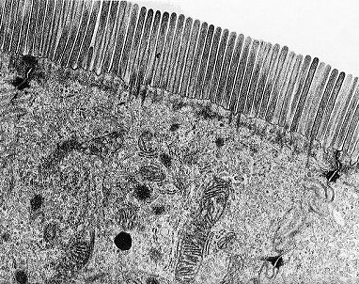 Microvilli at the cell surface
Microvilli at the cell surface
 Home
Home